
Dans cet article, je vais vous expliquer le sujet de mon mémoire, c’est-à-dire mon investigation pour déterminer si la mutation MDR1 influence le comportement chez le chien… et je vais en profiter pour ajouter un aspect “Philosophie des sciences”. Comme on va le voir, il y a plein de pistes, dans la recherche académique, qui ont été commencées, et puis sont finalement tombées en friche, sans raison apparentes. On va essayer de comprendre ce qui a pu se passer…
Des colleys décèdent pour des raisons mystérieuses…
Dans les années 80, des cas commencent à apparaitre, chez certains vétérinaires, de Colleys souffrant soudain de troubles neurologiques graves, menant même à la mort dans certains cas. Le point commun est souvent l’Ivermectine, un vermifuge régulièrement utilisé chez les chevaux et chez les chiens… mais pourquoi des affections neurologiques ? Et pourquoi seulement chez certains chiens, et de manière aussi grave, sans gradation ?

Des études montreront plus tard que l’Ivermectine, utilisée à des doses normales pour un usage comme vermifuge, ne devraient pas causer une telle réaction même chez ces chiens particuliers. Mais, fait peu connu, l’Ivermectine est aussi utilisée à l’époque en off label pour des affections cutanées… et là elle est utillisée à des doses largement supérieures à l’usage comme vermifuge. Les cas reportés ne mentionnent pas forcément la raison pour laquelle les chiens concernés ont reçu de l’Ivermectine, ni à quelle dose, puisqu’au moment où le vétérinaire est consulté, les troubles neurologiques sont là, et il est trop tard pour doser ce qui a été donné. Cette hypothèse expliquerait néanmoins la violence des symptômes, qui n’est pas forcément compatible avec les doses recommandées d’Ivermectine comme vermifuge, même chez ces chiens. On ne connait pas encore le mécanisme d’action à ce stade, mais il faut savoir que les symptômes neurologiques commencent à apparaitre à de petits multiplicateurs de la dose recommandées (3x, 4x la dose recommandée chez ces Colleys, alors que la dose létale chez les autres chiens est plus de 200x plus élevée): la marge thérapeutique est assez réduite, en particulier pour une espèce dont le poids peut énormément varier d’un individu à l’autre… En attendant que la recherche avance, le message est clair: ne pas utiliser l’Ivermectine chez les Colleys.
L’arrivée de Katrina Mealey, et la découverte du lien avec le gène MDR1

Un peu avant 2001 (date de sa publication Mealey et al., 2001 ), Katrina Mealey, vétérinaire et chercheuse à la Washington State University, a une de ces intuitions qui permettent à la recherche de faire des bonds…
Depuis les années 1970, la médecine est confrontée à un problème d’efficacité de certains médicaments anticancer: les médicaments ne parviennent pas à pénétrer suffisamment dans les organes visés. Le coupable est connu depuis au moins 1976 Schinkel et al., 1994 : il s’agit d’une protéine, la P-glycoprotéine, aussi appelée MDR1, qui protège certains organes (dont le cerveau) en évacuant des molécules spécifiques vers la circulation, les empêchant de pénétrer en quantité dans les organes concernés. Alfred Schinkel et son équipe au Netherlands Cancer Institute ont développé une lignée spécifique de souris ne disposant pas du gène permettant de produire cette protéine, à des fins de recherche. Et il s’avère que ces souris sont particulièrement sensibles… à l’Ivermectine.
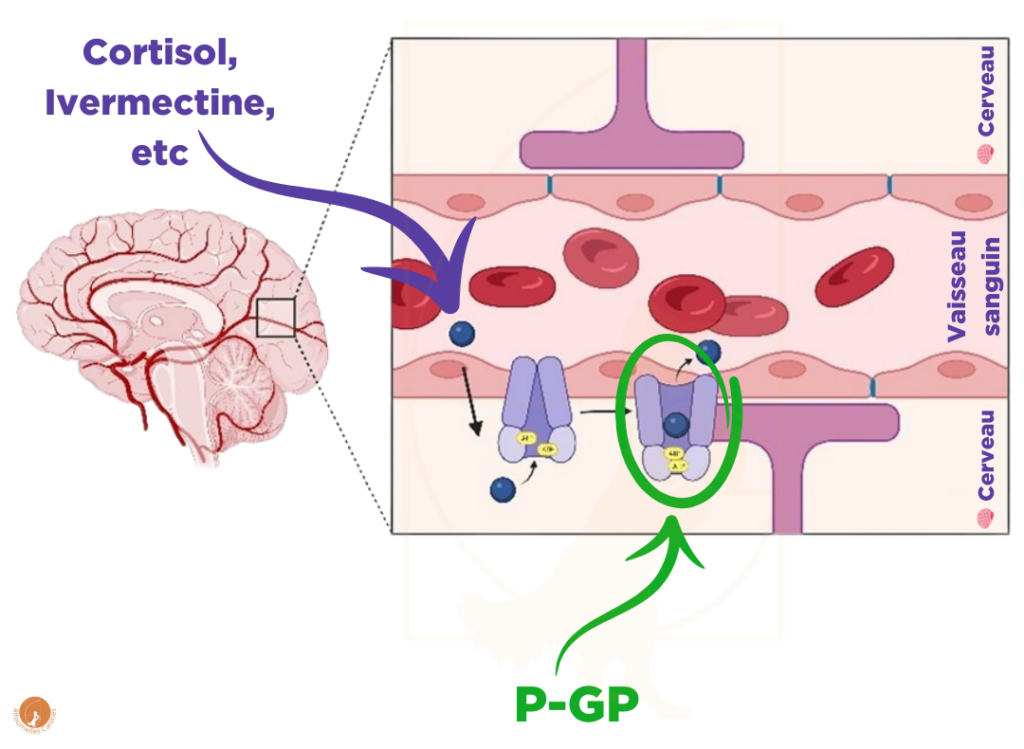
Katrina Mealey fait donc le lien, et teste cette hypothèse : elle mesure les taux d’Ivermectine dans le sang et dans le cerveau des Colleys décédés et des Beagles de laboratoire, séquence le gène MDR1 de ces chiens, et “Eurêka” ! Le lien est là ! Une mutation du gène MDR1 chez le chien serait donc responsable de cette hypersensibilité médicamenteuse qui cause des désordres neurologiques pouvant mener au décès.
A partir de là, la recherche peut tourner: on connait la cause de ce problème de médecine vétérinaire, et on a un modèle scientifique validé pour l’explorer. On peut tester tous les médicaments susceptibles d’être transportés par la P-gp grâce à des souris de laboratoire, et adapter les traitements pour les chiens. On peut aussi séquencer le gène MDR1 chez les chiens individuels susceptibles de porter la mutation, et on peut aussi mener des études sur les populations pour voir dans quelles races il est présent. Dans un sens, le problème est résolu : on est sur des rails…
Le lien entre P-gp et Cortisol
Mais à côté de ça, chez les humains, la recherche continue. Non seulement la P-gp reste un sujet de recherche important pour les traitements anticancer, mais il est également connu depuis pratiquement les débuts de la découverte de la P-gp que les stéroïdes, dont le Cortisol, font partie des molécules transportées par la P-gp…
Vous vous souvenez de l’article sur le Cortisol ? Au niveau de la barrière hémato-encéphalique se trouve la P-gp, qui raccompagne à la sortie une grande partie du Cortisol qui tente de pénétrer dans le cerveau. L’Hypothalamus jauge donc les besoins du corps en Cortisol sur un taux “filtré”, diminué par l’action de la P-gp.
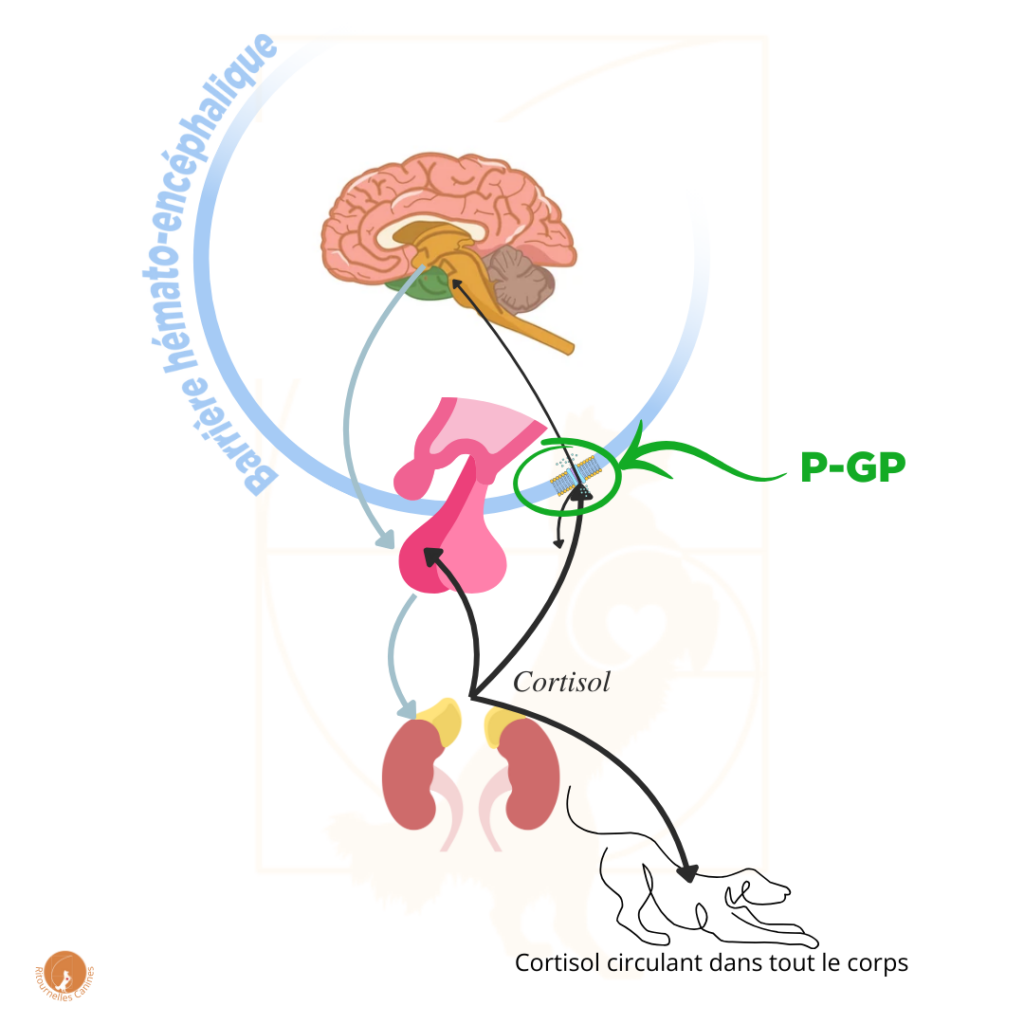
Or, chez les humains, une série de problématiques psychologiques sont liés au fonctionnement de l’axe du stress. Au-delà de la problématique vétérinaire canine, il y a donc encore des recherches qui se poursuivent sur la P-gp, et également sur son rôle au niveau de l’axe du stress.
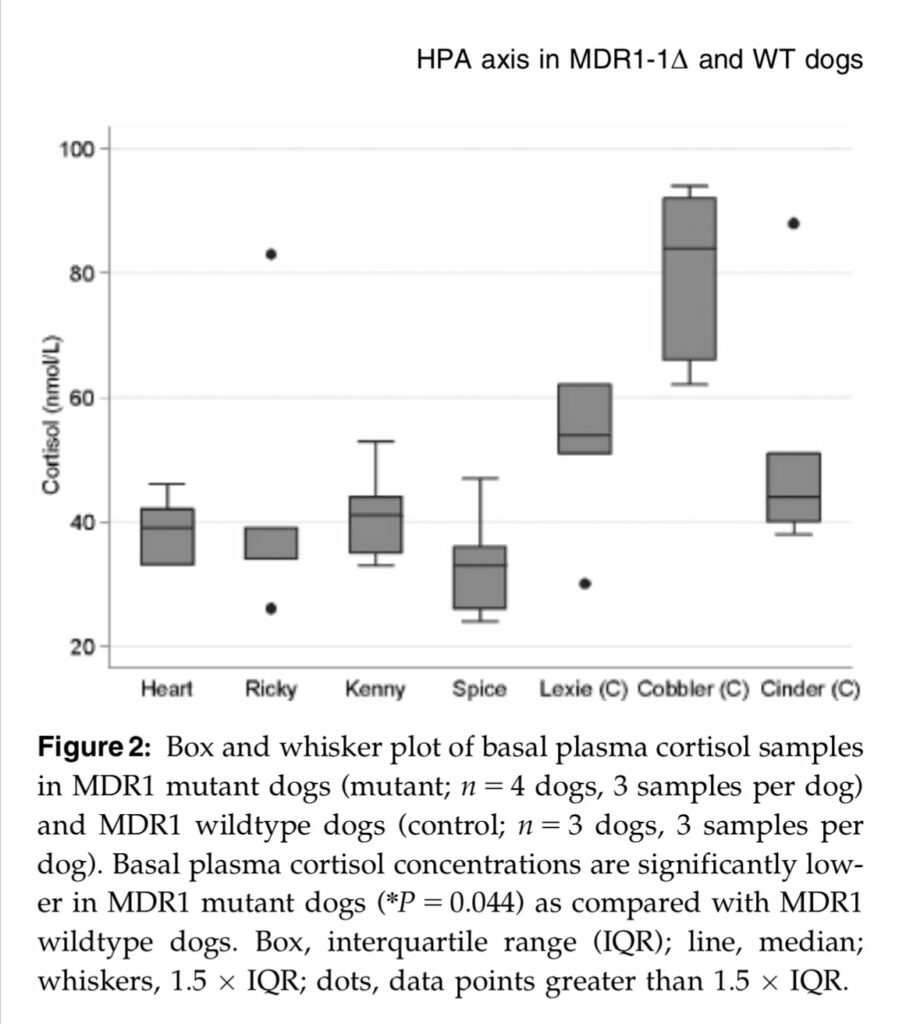
Une série de recherches sur les souris (et une chez les chiens… par Katrina Mealey et son équipe en 2007) montre notamment que les animaux dont la P-gp n’est pas fonctionnelle ont un profil particulier au niveau de l’axe du stress : vu que la P-gp ne fait pas son boulot de barrière, l’Hypothalamus reçoit plus de Cortisol que prévu. Or l’Hypothalamus ne sait pas qu’il y a un problème avec la P-gp et qu’elle doit adapter ses normes, évidemment. Donc elle envoie le signal de stopper la production de Cortisol alors que ce dernier n’est pas encore en niveaux suffisant pour le reste du corps. Résultat, on obtient un taux de Cortisol basal plus bas, et un pic de Cortisol lié au stress interrompu avant que tout le cycle du stress ait pu être complété (et accessoirement, les GR, eux, s’adaptent à ces taux bas en augmentant leur densité, ce qui rend les cellules encore plus sensibles aux moindres variations de Cortisol).
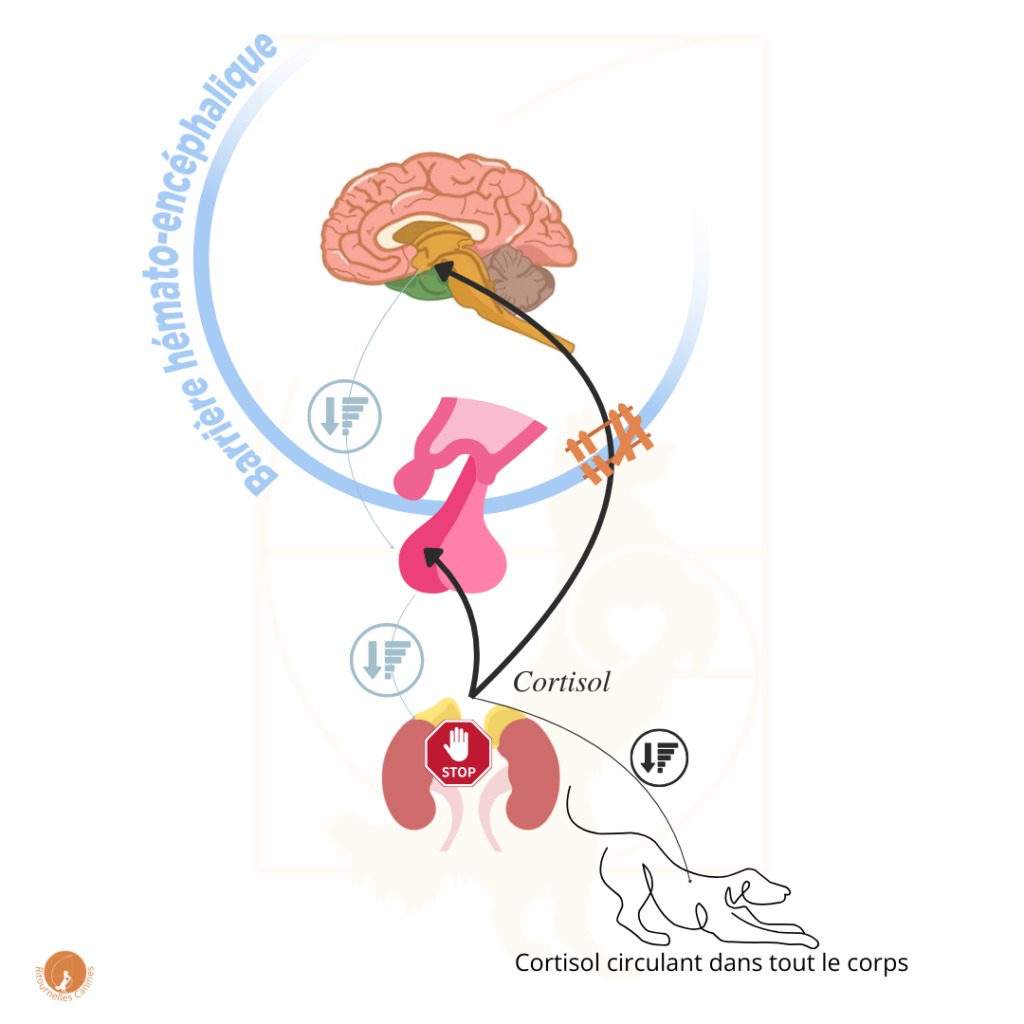
Curieusement, ce profil de fonctionnement de l’axe HPA, typique de la mutation MDR1, se retrouve également dans un autre profil psychologique, comme le montrent les travaux de Rachel Yehuda et son équipe : le stress post-traumatique (PTSD).
On retrouve donc, dans la recherche visant les humains, pas mal de développements concernant des profils de fonctionnement de l’axe HPA différents du fonctionnement normal, et qui sont corrélés à différentes problématiques psychologiques.
Pourtant, les recherches concernant le lien entre mutation MDR1 et Stress ne prennent pas… Pourquoi ?
En réalité, suite à ces découvertes du lien entre P-Gp, mutation, et problématiques comportementales notamment chez les souris, des recherches sont faites en termes de séquençage afin de voir si une telle mutation pourrait être corrélée à des problématiques psychologiques chez les humains. Or ces recherches ne donnent rien. Le sujet est donc plus ou moins abandonné, car il n’a pas d’intérêt pour le traitement des humains.
Et chez les chiens ?
Comme on l’a vu, le domaine principal de recherche visant la mutation MDR1 chez les chiens concerne le risque le plus important, car létal, c’est-à-dire la pharmacologie. Maintenant que cet aspect-là est plus ou moins réglé, il n’y a plus trop de raisons d’étendre le champ de recherche…
Du côté des pathologies liées au Cortisol, si on prend les symptômes cliniques vétérinaires (donc pas les symptômes comportementaux / psychologiques), la mutation n’a pas un impact suffisamment important par rapport aux pathologies bien connues comme la maladie d’Addison ou celle de Cushing. De plus, ces pathologies sont facilement identifiables sans devoir passer par la génétique : on voit s’il y a trop ou pas assez de Cortisol, ou voit si c’est lié à un problème de surrénales ou d’une tumeur hypophysaire, et on rééquilibre. La mutation altère l’axe HPA dans une limite qui n’est pas assez extrême pour faire apparaitre des symptômes physiologiques flagrants.
_Brussels%2C_BE.jpg)
Mais depuis une vingtaine d’années, la cognition canine devient un important domaine de recherche, lié notamment à un changement de perspective en termes de valeur dans notre rapport à l’animal. Pour caricaturer, avant, le chien était vu comme un animal bâtard, indigne de l’intérêt des éthologues car pas assez “naturel”, trop domestiqué, et pas très intéressant non plus pour les sciences cognitives car il n’apportait pas grand chose, coincé entre les humains (le top, mais on ne peut pas faire d’expériences invasives dessus), les grands singes (”proches” de l’humain sans être l’humain, donc prometteurs) et les rongeurs (sur lesquels on peut faire les expériences qu’on veut). Aujourd’hui, le chien est vu comme justement rendu particulièrement intéressant par son statut d’animal “bâtard”, au croisement entre la “nature” et la “civilisation” humaine: un animal qui vit dans le même monde urbanisé que nous, dans nos maisons, mais qui en même temps n’est pas l’humain.
Bref, il y avait donc là un vide, parce que la recherche sur le MDR1 était passée à autre chose, malgré que le chemin était déjà en grande partie pavé. Et pourtant, dans le même temps, on commence à s’intéresser de plus en plus à l’influence des pathologies de l’axe du stress sur le comportement canin, comme en témoignent les recherches de l’équipe de Daniel Mills…
Mon sujet de recherche
Récapitulons. Actuellement, avec ce qu’on a vu, au niveau de la recherche, on en est là:
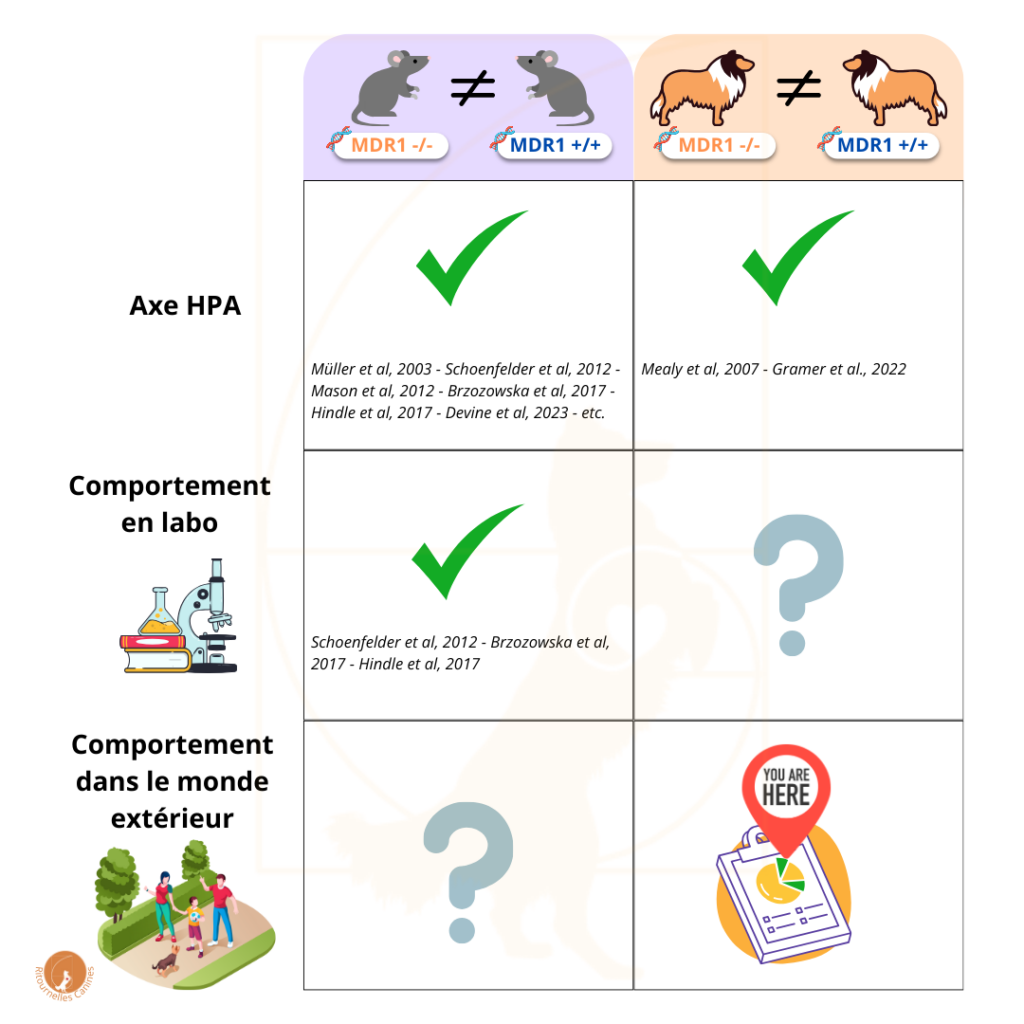
Le lien entre la mutation MDR1 et l’axe HPA a été démontré chez les souris et chez les chiens, et l’impact sur le comportement a été démontré chez des souris de laboratoire. Il reste encore à déterminer si l’impact sur le comportement canin se vérifie…
En théorie, pour faire une étude objective, il faudrait pouvoir mesurer tout ça en laboratoire. Malheureusement, d’une part je n’avais pas accès à ce type d’infrastructure (et de financement), mais d’autre part, il y a aussi une raison plus fondamentale qui rendait cette méthode inadéquate : pour pouvoir tester le comportement de chiens en laboratoire, encore faut-il que ces chiens parviennent à se comporter normalement dans le cadre de cette infrastructure, pour que le test puisse avoir lieu dans des bonnes conditions. Or, dans ce cas-ci, vu que l’hypothèse est justement que, si cela a un impact, cet impact porte justement sur leur résilience et leur capacité à gérer le stress et la nouveauté… ces chiens ne se comporteraient probablement pas normalement dans le contexte d’un laboratoire !
Je devais donc recourir à l’autre design de recherche couramment utilisé en comportement canin, à savoir une recherche par questionnaire. Evidemment, cela amène d’emblé des limitations:
- Il peut y avoir des divergences d’interprétation d’une personne à l’autre (par exemple, prenons une question demandant si le chien aboie souvent, de 1 à 5. Pour une personne, si son chien aboie une fois par jour, c’est “souvent”, et il mettra 4, alors que pour une autre personne, “souvent” c’est toutes les heures, donc cette personne mettra 1 si son chien aboie une fois par jour).
- On ne touchera que des personnes déjà engagées sur le sujet. Par exemple, sur un groupe lié à la réactivité canine, on aura plus de gens dont les chiens sont réactifs que dans la population générale, et probablement des gens qui sont en démarche de recherche de solution
- Ce sont des chiens qui vivent dans le monde extérieur, donc il y a une variabilité et de multiples facteurs non contrôlés qui interviennent. Ce sont forcément tous des chiens qui ont des vécus différents dont on ne connait rien, qui vivent dans des situations toutes différentes, avec des gardien·ne·s qui n’ont pas tou·te·s le même rapport au chien ni même les mêmes compétences de lecture de leur chien.
Au final, même en ayant bien construit le questionnaire, il est fort probable que la différence que fait la mutation soit noyée dans toute la variabilité des autres facteurs. Peut-être qu’il y a une différence, mais qu’elle passera sous les radars de mon étude… C’est un risque à prendre.
La méthode
J’ai commencé par relever dans la littérature académique tous les comportements qui avaient été relevés comme étant corrélés à des différences de taux de cortisol ou de fonctionnement de l’axe HPA. Vu qu’il y a assez peu d’études là-dessus chez les chiens, j’ai également regardé ce que la recherche indiquait pour les rongeurs et pour les humains, pour voir si on pouvait prudemment faire des transpositions… mais pas seulement.
En analysant tout cela, j’ai tenté d’identifier:
- d’une part, des différences spécifiques à ce profil de fonctionnement de l’axe HPA (bas cortisol basal, boucle de feedback négatif accrue, plus grande densité de GR), donc pas les comportements qui apparaissent dans d’autres profils.
- d’autre part, des différences non-spécifiques à un profil, mais qui témoignent quand même d’une altération de l’axe HPA. Par exemple, une instabilité d’humeur ou de l’anxiété peuvent apparaitre dans plusieurs profils différents voire contradictoires (cortisol trop bas mais aussi cortisol trop haut) : ils sont donc symptômatiques d’une altération de l’axe HPA de manière générale.
A partir de là, j’ai pu construire un formulaire qui portait sur des comportements susceptibles d’être différents entre un chien avec un axe HPA normal, et un chien qui porterait la mutation.
Au final je suis arrivé aux questions suivantes:
(insérer les questions)
Evidemment, rien n’était souligné en gras dans le questionnaire soumis, j’ai ajouté cela ici pour la facilité de lecture… De même, je n’encourage certainement pas de réprimander son chien, mais cette question était pertinente dans ce cadre, parce qu’il y a des gardien·ne·s qui le font, et la réaction du chien peut être symptômatique (cette question vient d’un questionnaire de l’équipe de Daniel Mills). Ceux et celles d’entre vous qui sont familier·e·s avec des chiens sensibles, anxieux, réactifs, reconnaitront pas mal de traits: ils ne sont pas forcément spécifiques à la mutation, mais ils sont pertinents pour comparer les chiens qui possèdent la mutation par rapport à ceux qui ne la possèdent pas: si ceux qui ont la mutation scorent statistiquement plus haut, en moyenne, sur certaines questions, que les chiens sans mutation, il est fort probable que ce soit lié à la mutation, sinon les moyennes ne seraient pas différentes.
J’ai ensuite soumis le questionnaire à un maximum de groupes Facebook centrés sur les Colleys, pour limiter au moins le facteur race, et j’ai analysé les résultats au moyen de méthodes statistiques. D’ailleurs, c’est assez intéressant de noter que même dans ce domaine (et c’est logique puisque c’est un domaine scientifique), il existe des débats et donc des positions différentes sur les méthodes les plus adaptées à utiliser dans tel ou tel cas… C’est loin d’être aussi clair qu’il n’y parait !
Résultats
Au final, j’ai obtenu 140 réponses, dont 108 Colleys dont le statut MDR1 était connu par leur gardien·ne·s. J’ai procédé à différentes analyses (le fameux χ2), de la plus large à la plus précise (le tau-c de Kendall), pour voir ce qui ressortait. Il existe une accusation, dans le monde de l’analyse statistique, qui est celle de faire du “p-hacking”, c’est-à-dire d’appliquer plein de tests différents un peu comme on va à la pêche, en espérant tomber à force sur un résultat significatif. On est donc tenus de prouver que ce n’est pas ce que l’on fait, de montrer patte blanche, de différentes façons. Or, il y a aussi, comme dans tous les domaines académiques, une obligation implicite : celle d’utiliser des méthodes acceptées par les collègues, peu importe qu’elles soient les plus adaptées ou non. Par exemple, dans les études de comportement canin, il est plus facile d’utiliser le C-BARQ, qui est le test de comportement le plus reconnu, que d’aller créer un nouveau questionnaire. On va parfois donc utiliser une méthode qu’on trouve problématique (par exemple, le C-BARQ associe certains comportements à la peur, et d’autres à l’agression, qui deviennent deux traits exclusifs : c’est soit l’un, soit l’autre. Et un chien qui aboie est donc agressif, et pas peureux), parce que ça évitera des problèmes avec les collègues… ou dans le cas des publications peer-review, avec les reviewers. Il n’est pas rare en effet que les reviewers critiquent une étude parce que l’auteur n’a pas utilisé la méthode classique, et qu’ils ne veulent pas chercher plus loin, il faut utiliser la méthode classique. Vu l’investissement en temps et en argent que demandent les publications, cela encourage à utiliser les méthodes qui auront le plus de chances de plaire aux reviewers plutôt que les méthodes les plus adaptées. Il y a également un autre non-dit : plus les études utilisent les mêmes méthodes, plus il est facile des les comparer avec des méta-analyses, aussi critiquable que soit cette technique qui broie les spécificités des études sur l’autel des moyennes. Bref pas étonnant que tout pousse à utiliser les mêmes méthodes, peu importe si elles sont les plus adaptées ou non…
Dans ce cas-ci, au final, c’est un peu ce que j’ai fait : après quelques recherches, la méthode d’analyse la plus adaptée pour mes données était le tau-c de Stuart-Kendall. On s’en fiche, mais ce qui est intéressant, est que cette méthode n’est pas prévue en standard dans beaucoup de logiciels : c’est dire si elle est spéficique. Donc pour éviter les accusations de p-hacking et donc d’avoir été chercher une méthode tordue et inconnue pour arriver à avoir des résultats, j’ai également inclus d’autres méthodes comme le fameux χ2, qui est la méthode la plus utilisée dans les études de comportement canin, et deux autres méthodes intermédiaires, plus précises mais aussi moins utilisées. Je ne rentre pas dans les détails, mais je trouvais intéressant de saisir ce point pour développer un aspect intéressant de Philosophie des Sciences relatif aux méthodes statistiques.
Bref, au final, quelques corrélations significatives sont apparues:
Q2. Certains chiens réagissent très fortement (aboiements, etc) lorsqu’ils sont surpris, ne réalisant qu’après coup que ce qui les a surpris était en réalité inoffensif ou bien connu d’eux, tandis que d’autres chiens lèvent à peine une oreille et réfléchissent avant de décider si cela vaut la peine de réagir. Sur une échelle de 1 à 7, où 1 correspond à « même surpris, observe avant d’agir » et 7 à « quand il est surpris, réagit immédiatement de façon indiscriminée« , comment définiriez-vous le comportement de votre chien ?
Q3. Certains chiens ont tendance à sursauter très facilement, par exemple lorsqu’ils voient ou entendent quelque chose de soudain ou d’inhabituel. Sur une échelle de 1 à 7, où 1 correspond à un « sursaute rarement / est difficilement surpris » et 7 à une « se surprend facilement / réagit de manière excessive lorsqu’il est surpris », comment définiriez-vous le comportement de votre chien ?
Sur ces deux questions, le test du χ2, c’est-à-dire le test le plus global, indique une corrélation significative entre les chiens affectés par la mutation est les chiens non-affectés.
De manière intéressante, lorsqu’on utilise les méthodes plus fines (le tau-c et Kruskall-Wallis), une corrélation significative apparait sur une autre question:
Q15. Certains chiens semblent ne pouvoir se relaxer que dans des environnements extrêmement calmes. Ils sont particulièrement perceptifs, et même des stimuli visuels ou auditifs de faible intensité comme de la musique, la TV, ou le chat qui court dans la pièce, les dérangent. Sur une échelle de 1 à 7, où 1 correspond à “il peut dormir dans n’importe quel environnement, peu importe le bruit ou les distractions visuelles” et 7 à “il a besoin du silence total dans la pièce pour arriver à se relaxer complètement”, comment évaluez-vous le comportement de votre chien ?
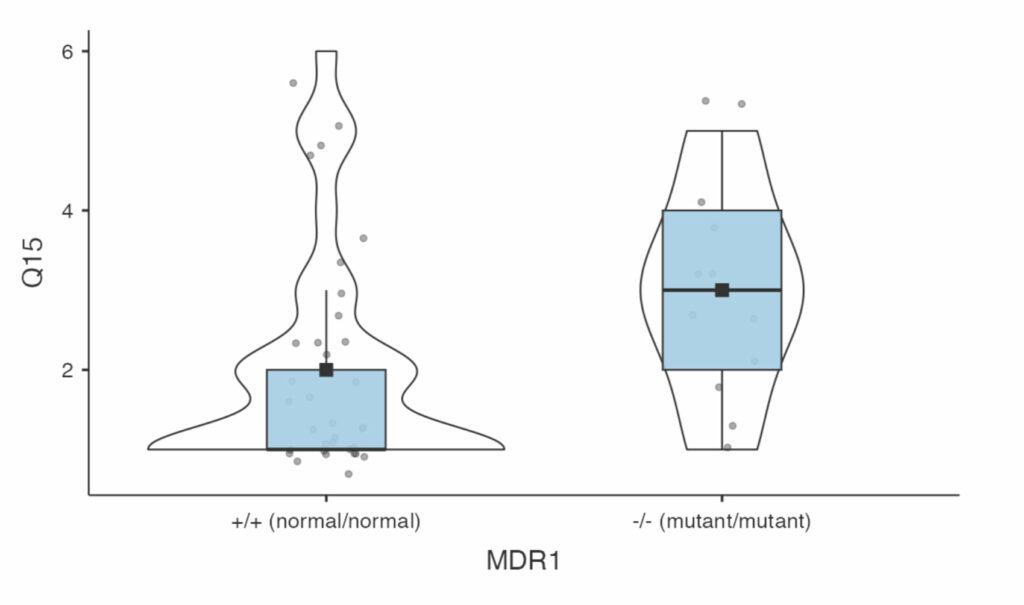
A ce stade, il faut noter que dans ces premières analyses, j’ai été beaucoup plus large que les auteurs des études sur les souris: eux n’ont inclus que les cas extrêmes (les souris complètement indemnes et les souris qui portent deux copies de la mutation, en excluant les souris qui portent un gène normal et un gène mutant), alors que moi j’avais inclus les chiens “+/-”, car ils sont certainement affectés dans dans une moindre mesure.
Par souci de comparaison, j’ai ensuite utilisé les mêmes critères qu’eux, donc j’ai exclu les chiens “+/-” pour ne garder que les cas extrêmes. Et là, sans surprise, Q3 et Q15 montrent une corrélation significative entre score sur cette question et statut MDR1.
Q11. Certains chiens ont une tendance marquée à éviter les personnes ou les situations inconnues ou non familières, par exemple ils ont tendance à quitter la pièce lorsque des invités inconnus arrivent ou lorsque des personnes crient ou qu’il y a des bruits forts. Sur une échelle de 1 à 7, où 1 correspond à « aucune tendance à se cacher ou à éviter les gens ou les situations » et 7 à « forte tendance à se cacher ou à éviter les gens ou les situations », comment évaluez-vous le comportement de votre chien ?
A noter qu’une autre question arrive tout juste sous le seuil de significativité de p < 0,05 :
Q9. En pensant au tempérament de votre chien, comment définiriez-vous son niveau de nervosité/agitation sur une échelle de 1 à 7 où 1 correspond à “très calme” et 7 à « très nerveux et agité » ?
Je vous passe ici plein de détails sur lesquels je veux bien discuter ailleurs, avec plaisir, mais ça ne vaut pas la peine de rallonger l’article pour tout le monde avec ça…
D’accord, mais qu’est-ce qu’on fait avec ça ?
Globalement, il faudrait plus d’études sur le sujet pour voir si ces résultats se confirment ou non, il faudrait aussi faire la même étude pour d’autres races comme par exemple le Berger Australien, mais ces résultats permettent en tout cas de supposer qu’au-delà de tous les autres facteurs individuels, l’altération de l’axe du stress causée par la mutation MDR1 pourrait effectivement bien avoir un impact sur le caractère de nos chiens, les rendant plus susceptibles d’être particulièrement sensibles aux stimuli inattendus, plus vulnérables à l’anxiété et aux stimulations sensorielles, qu’ils auraient besoin de plus de calme et de silence pour arriver à réellement récupérer, et qu’ils seraient plus méfiants vis-à-vis de la nouveauté.
Fondamentalement, être attentifs à la qualité de l’environnement de repos de nos chiens (voir le Sanctuaire), être vigilants à leur résilience et à leur capacité à appréhender la nouveauté (voir la Ritournelle), et à leur capacité à gérer les contrastes environnementaux, ce sont des conseils de prudence qui sont bénéfiques pour tous les chiens. Mais il est possible que les chiens porteurs du gène MDR1 aient des prédispositions à être particulièrement sensibles sur ces points, et qu’il serait intéressant d’être encore plus attentifs à cela avec eux… Mais comme le dit la formule : “further research is needed” !
LA référence en Français sur le MDR1, avec la liste des médicaments à garder à l’oeil :
Une excellente synthèse par l’équipe de Katrina Mealey sur l’état des connaissances scientifiques actuelles sur le MDR1/abcb1 chez le chien et le chat :
Un super article de l’équipe de Daniel Mills sur les symptômes comportementaux chez les chiens traités avec de la cortisone :
Un livre indispensable sur le stress et le cortisol chez les chiens :

Amat, M., Le Brech, S. and Manteca, X. (2024) ‘The Relationship Between Aggression and Physical Disease in Dogs’, The Veterinary Clinics of North America. Small Animal Practice, 54(1), pp. 43–53. Available at: https://doi.org/10.1016/j.cvsm.2023.08.008.
Arnett, M.G. et al. (2016) ‘Genetic Approaches to Hypothalamic-Pituitary-Adrenal Axis Regulation’, Neuropsychopharmacology, 41(1), pp. 245–260. Available at: https://doi.org/10.1038/npp.2015.215.
Arvelius, P. et al. (2014) ‘Genetic analysis of a temperament test as a tool to select against everyday life fearfulness in Rough Collie1’, Journal of Animal Science, 92(11), pp. 4843–4855. Available at: https://doi.org/10.2527/jas.2014-8169.
Arvelius, P. (2015) ‘Genetic evaluation of behaviour in dogs’, Journal of Veterinary Behavior, 10(5), p. 439. Available at: https://doi.org/10.1016/j.jveb.2015.07.004.
Baars, C. et al. (2008) ‘Allele-specific polymerase chain reaction diagnostic test for the functional MDR1 polymorphism in dogs’, The Veterinary Journal, 177(3), pp. 394–397. Available at: https://doi.org/10.1016/j.tvjl.2007.05.020.
Baker, M.E. and Katsu, Y. (2019) ‘Chapter Two – Evolution of the Mineralocorticoid Receptor’, in G. Litwack (ed.) Vitamins and Hormones. Academic Press (Aldosterone), pp. 17–36. Available at: https://doi.org/10.1016/bs.vh.2018.10.009.
Barroso, M.C. et al. (2022) ‘Occurrence of MDR1 1-delta mutation in herding dog breeds in Portugal’, Frontiers in Veterinary Science, 9, p. 990884. Available at: https://doi.org/10.3389/fvets.2022.990884.
Beckers, E. et al. (2022) ‘The prevalence of the ABCB1-1Δ variant in a clinical veterinary setting: The risk of not genotyping’, PLoS ONE, 17(8), p. e0273706. Available at: https://doi.org/10.1371/journal.pone.0273706.
Bell Rizzolo, J. and Bradshaw, G.A. (2016) ‘Prevalence and Patterns of Complex PTSD in Asian Elephants (Elephas maximus)’, in, pp. 291–297.
Bloise, E. and Matthews, S.G. (2019) ‘Multidrug Resistance P-Glycoprotein (P-gp), Glucocorticoids, and the Stress Response’, in Stress: Physiology, Biochemistry, and Pathology. Elsevier, pp. 227–241. Available at: https://doi.org/10.1016/B978-0-12-813146-6.00019-9.
Braem Dubé, M. et al. (2020) ‘Parallels in the interactive effect of highly sensitive personality and social factors on behaviour problems in dogs and humans’, Scientific Reports, 10(1), p. 5288. Available at: https://doi.org/10.1038/s41598-020-62094-9.
Braem, M. et al. (2017) ‘Development of the “Highly Sensitive Dog” questionnaire to evaluate the personality dimension “Sensory Processing Sensitivity” in dogs’, PLOS ONE. Edited by V. Miragliotta, 12(5), p. e0177616. Available at: https://doi.org/10.1371/journal.pone.0177616.
Braem, M. (2024) ‘Sensory Processing Sensitivity and the Importance of Individuality and Personality in Veterinary Medicine’, Veterinary Clinics of North America: Small Animal Practice, 54(1), pp. 181–193. Available at: https://doi.org/10.1016/j.cvsm.2023.09.002.
Bremner, J.D. (1999) ‘Does stress damage the brain?’, Biological Psychiatry, 45(7), pp. 797–805. Available at: https://doi.org/10.1016/S0006-3223(99)00009-8.
Brzozowska, N.I. et al. (2017) ‘Genetic deletion of P-glycoprotein alters stress responsivity and increases depression-like behavior, social withdrawal and microglial activation in the hippocampus of female mice’, Brain, Behavior, and Immunity, 65, pp. 251–261. Available at: https://doi.org/10.1016/j.bbi.2017.05.008.
Can, A., Dao, D.T., Arad, M., et al. (2012) ‘The Mouse Forced Swim Test’, Journal of Visualized Experiments : JoVE, (59), p. 3638. Available at: https://doi.org/10.3791/3638.
Can, A., Dao, D.T., Terrillion, C.E., et al. (2012) ‘The Tail Suspension Test’, Journal of Visualized Experiments : JoVE, (59), p. 3769. Available at: https://doi.org/10.3791/3769.
Cavigelli, S.A. et al. (2007) ‘Behavioral inhibition and glucocorticoid dynamics in a rodent model’, Physiology & Behavior, 92(5), pp. 897–905. Available at: https://doi.org/10.1016/j.physbeh.2007.06.016.
Chakraborty, P., Chattarji, S. and Jeanneteau, F. (2021) ‘A salience hypothesis of stress in PTSD’, European Journal of Neuroscience, 54(11), pp. 8029–8051. Available at: https://doi.org/10.1111/ejn.15526.
Chattarji, S. and Rao, R.P. (2014) ‘Blood–brain biomarkers for stress susceptibility’, Proceedings of the National Academy of Sciences, 111(37), pp. 13253–13254. Available at: https://doi.org/10.1073/pnas.1414663111.
Cocco, R. et al. (2023) ‘A Preliminary Study on the Interplay between the Serum Levels of Neurotransmitters and Thyroid Hormones for the Evaluation of the Behavioral Phenotype of Dogs’, Animals, 13(3), p. 411. Available at: https://doi.org/10.3390/ani13030411.
Cohen, H., Matar, M.A. and Zohar, J. (2014) ‘Maintaining the Clinical Relevance of Animal Models in Translational Studies of Post-Traumatic Stress Disorder’, ILAR Journal, 55(2), pp. 233–245. Available at: https://doi.org/10.1093/ilar/ilu006.
Corridan, C.L., Dawson, S.E. and Mullan, S. (2024) ‘Potential Benefits of a “Trauma-Informed Care” Approach to Improve the Assessment and Management of Dogs Presented with Anxiety Disorders’, Animals, 14(3), p. 459. Available at: https://doi.org/10.3390/ani14030459.
Daskalakis, N.P. et al. (2014) ‘Expression profiling associates blood and brain glucocorticoid receptor signaling with trauma-related individual differences in both sexes’, Proceedings of the National Academy of Sciences, 111(37), pp. 13529–13534. Available at: https://doi.org/10.1073/pnas.1401660111.
Daskalakis, N.P. et al. (2016) ‘Endocrine Aspects of PTSD: Hypothalamic-Pituitary-Adrenal (HPA) Axis and Beyond’, in C.R. Martin, V.R. Preedy, and V.B. Patel (eds) Comprehensive Guide to Post-Traumatic Stress Disorders. Cham: Springer International Publishing, pp. 245–260. Available at: https://doi.org/10.1007/978-3-319-08359-9_130.
Daskalakis, N.P. et al. (2022) ‘Mineralocorticoid receptor and glucocorticoid receptor work alone and together in cell-type-specific manner: Implications for resilience prediction and targeted therapy’, Neurobiology of Stress, pp. 100455–100455. Available at: https://doi.org/10.1016/j.ynstr.2022.100455.
De Kloet, E.R. (2023) ‘Glucocorticoid feedback paradox: a homage to Mary Dallman’, Stress, 26(1), p. 2247090. Available at: https://doi.org/10.1080/10253890.2023.2247090.
De Kloet, E.R. and Joëls, M. (2023) ‘The cortisol switch between vulnerability and resilience’, Molecular Psychiatry [Preprint]. Available at: https://doi.org/10.1038/s41380-022-01934-8.
Dekel, Y. et al. (2017) ‘Frequency of canine nt230(del4) MDR1 mutation in prone pure breeds, their crosses and mongrels in Israel – insights from a worldwide comparative perspective’, BMC veterinary research, 13(1), p. 333. Available at: https://doi.org/10.1186/s12917-017-1251-9.
Department of Genetics and Animal Breeding, Warsaw University of Life Sciences, Poland et al. (2017) ‘DRUG TOLERANCE/INTOLERANCE IN DOMESTIC DOG (Canis lupus familiaris) WITH MDR1 GENE DEFECT’, Folia Pomeranae Universitatis Technologiae Stetinensis Agricultura, Alimentaria, Piscaria et Zootechnica, 332(41), pp. 117–126. Available at: https://doi.org/10.21005/AAPZ2017.41.1.12.
Devine, K. et al. (2023) ‘The ATP-binding cassette proteins ABCB1 and ABCC1 as modulators of glucocorticoid action’, Nature Reviews Endocrinology, 19(2), pp. 112–124. Available at: https://doi.org/10.1038/s41574-022-00745-9.
Dissertation (no date) Google Docs. Available at: https://docs.google.com/document/d/1QS-JSwpv5Khr9NM_Vw5sxFL4kRezQWcKQ5amEDwvXp4/edit?usp=drive_web&ouid=108265496567630362295&usp=embed_facebook (Accessed: 21 November 2024).
Eken Asp, H. (2015) Everyday behaviour in dogs. Available at: https://res.slu.se/id/publ/68352 (Accessed: 21 June 2024).
Engel, S. et al. (2023) ‘Cortisol response to traumatic stress to predict PTSD symptom development – a systematic review and meta-analysis of experimental studies’, European Journal of Psychotraumatology, 14(2), p. 2225153. Available at: https://doi.org/10.1080/20008066.2023.2225153.
Engel-Yeger, B., Palgy-Levin, D. and Lev-Wiesel, R. (2013) ‘The Sensory Profile of People With Post-Traumatic Stress Symptoms’, Occupational Therapy in Mental Health, 29(3), pp. 266–278. Available at: https://doi.org/10.1080/0164212X.2013.819466.
Enokido, M. et al. (2014) ‘Implication of P-Glycoprotein in Formation of Depression-Prone Personality: Association Study between the C3435T MDR1 Gene Polymorphism and Interpersonal Sensitivity’, Neuropsychobiology, 69(2), pp. 89–94. Available at: https://doi.org/10.1159/000358063.
Erkens, T. et al. (2009) ‘Presence of the ABCB1 (MDR1) deletion mutation causing ivermectin hypersensitivity in certain dog breeds in Belgium’, (2009) VLAAMS DIERGENEESKUNDIG TIJDSCHRIFT [Preprint]. Available at: http://lib.ugent.be/catalog/pug01:740545.
Fan, Z. et al. (2021) ‘Environmental enrichment modulates HPA axis reprogramming in adult male rats exposed to early adolescent stress’, Neuroscience Research, 172, pp. 63–72. Available at: https://doi.org/10.1016/j.neures.2021.04.007.
Fatjó, J. and Bowen, J. (2020) ‘Making the Case for Multi-Axis Assessment of Behavioural Problems’, Animals, 10(3), p. 383. Available at: https://doi.org/10.3390/ani10030383.
Fenster, R.J. et al. (2018) ‘Brain circuit dysfunction in post-traumatic stress disorder: from mouse to man’, Nature Reviews Neuroscience, 19(9), pp. 535–551. Available at: https://doi.org/10.1038/s41583-018-0039-7.
Firdova, Z. et al. (2016) ‘The prevalence of ABCB1:c.227_230delATAG mutation in affected dog breeds from European countries’, Research in Veterinary Science, 106, pp. 89–92. Available at: https://doi.org/10.1016/j.rvsc.2016.03.016.
Fleming, L.L., Harnett, N.G. and Ressler, K.J. (2024) ‘Sensory alterations in post-traumatic stress disorder’, Current Opinion in Neurobiology, 84, p. 102821. Available at: https://doi.org/10.1016/j.conb.2023.102821.
Flory, J.D. and Yehuda, R. (2017) ‘Chapter 30 – Glucocorticoid Receptor: Genetics and Epigenetics in Veterans With PTSD’, in G. Fink (ed.) Stress: Neuroendocrinology and Neurobiology. San Diego: Academic Press, pp. 303–307. Available at: https://doi.org/10.1016/B978-0-12-802175-0.00030-9.
Fries, E. et al. (2005) ‘A new view on hypocortisolism’, Psychoneuroendocrinology, 30(10), pp. 1010–1016. Available at: https://doi.org/10.1016/j.psyneuen.2005.04.006.
Fujii, T. et al. (2012) ‘Association between the functional polymorphism (C3435T) of the gene encoding P-glycoprotein (ABCB1) and major depressive disorder in the Japanese population’, Journal of Psychiatric Research, 46(4), pp. 555–559. Available at: https://doi.org/10.1016/j.jpsychires.2012.01.012.
Geyer, J. et al. (2005) ‘Development of a PCR-based diagnostic test detecting a nt230(del4) MDR1 mutation in dogs: verification in a moxidectin-sensitive Australian Shepherd’, Journal of Veterinary Pharmacology and Therapeutics, 28(1), pp. 95–99. Available at: https://doi.org/10.1111/j.1365-2885.2004.00625.x.
Gramer, I. et al. (2011) ‘Breed distribution of the nt230(del4) MDR1 mutation in dogs’, The Veterinary Journal, 189(1), pp. 67–71. Available at: https://doi.org/10.1016/j.tvjl.2010.06.012.
Gramer, I. et al. (2022) ‘Urinary cortisol metabolites are reduced in MDR1 mutant dogs in a pilot targeted GC‐MS urinary steroid hormone metabolome analysis’, Journal of Veterinary Pharmacology and Therapeutics, 45(3), pp. 265–272. Available at: https://doi.org/10.1111/jvp.13050.
Guzmán Ramos, P.J. et al. (2022) ‘Diagnosis of canine spontaneous hypoadrenocorticism’, Canine Medicine and Genetics, 9(1), p. 6. Available at: https://doi.org/10.1186/s40575-022-00119-4.
Harricharan, S., McKinnon, M.C. and Lanius, R.A. (2021) ‘How Processing of Sensory Information From the Internal and External Worlds Shape the Perception and Engagement With the World in the Aftermath of Trauma: Implications for PTSD’, Frontiers in Neuroscience, 15. Available at: https://doi.org/10.3389/fnins.2021.625490.
Henckens, M.J.A.G. et al. (2016) ‘Interindividual differences in stress sensitivity: basal and stress-induced cortisol levels differentially predict neural vigilance processing under stress’, Social Cognitive and Affective Neuroscience, 11(4), pp. 663–673. Available at: https://doi.org/10.1093/scan/nsv149.
Hindle, S.J. et al. (2017) ‘Evolutionarily Conserved Roles for Blood-Brain Barrier Xenobiotic Transporters in Endogenous Steroid Partitioning and Behavior’, Cell Reports, 21(5), pp. 1304–1316. Available at: https://doi.org/10.1016/j.celrep.2017.10.026.
Horn, S.R. and Feder, A. (2018) ‘Understanding Resilience and Preventing and Treating PTSD’, Harvard Review of Psychiatry, 26(3), pp. 158–174. Available at: https://doi.org/10.1097/HRP.0000000000000194.
Horváth, Z. et al. (2007) ‘Three different coping styles in police dogs exposed to a short-term challenge’, Hormones and Behavior, 52(5), pp. 621–630. Available at: https://doi.org/10.1016/j.yhbeh.2007.08.001.
Hugnet, C., Bentjen, S.A. and Mealey, K.L. (2004) ‘Frequency of the mutant MDR1 allele associated with multidrug sensitivity in a sample of collies from France’, Journal of Veterinary Pharmacology and Therapeutics, 27(4), pp. 227–229. Available at: https://doi.org/10.1111/j.1365-2885.2004.00585.x.
Joëls, M. (2006) ‘Corticosteroid effects in the brain: U-shape it’, Trends in Pharmacological Sciences, 27(5), pp. 244–250. Available at: https://doi.org/10.1016/j.tips.2006.03.007.
Karssen, A.M. et al. (2001) ‘Multidrug Resistance P-Glycoprotein Hampers the Access of Cortisol But Not of Corticosterone to Mouse and Human Brain’, Endocrinology, 142(6), pp. 2686–2694. Available at: https://doi.org/10.1210/endo.142.6.8213.
Kawabata, A. et al. (2005) ‘Canine mdr1 Gene Mutation in Japan’, Journal of Veterinary Medical Science, 67(11), pp. 1103–1107. Available at: https://doi.org/10.1292/jvms.67.1103.
Kintzer, P.P. and Peterson, M.E. (1997) ‘Primary and Secondary Canine Hypoadrenocorticism’, Veterinary Clinics of North America: Small Animal Practice, 27(2), pp. 349–357. Available at: https://doi.org/10.1016/S0195-5616(97)50036-2.
Klein, S.C. and Peterson, M.E. (2010a) ‘Canine hypoadrenocorticism: Part I’, The Canadian Veterinary Journal, 51(1), pp. 63–69.
Klein, S.C. and Peterson, M.E. (2010b) ‘Canine hypoadrenocorticism: Part II’, The Canadian Veterinary Journal, 51(2), p. 179.
Klerk, O.L. de (2011) The guarded brain: the role of P-glycoprotein at the blood-brain barrier in major psychiatric disorders and antidepressant treatment. Groningen: s.n.
de Kloet, E.R. (2024) ‘Coping with the multifaceted and multifunctional role of cortisol in the brain’, Neuroscience Applied, 3, p. 104047. Available at: https://doi.org/10.1016/j.nsa.2024.104047.
Lensen, R.C.M.M., Moons, C.P.H. and Diederich, C. (2019) ‘Physiological stress reactivity and recovery related to behavioral traits in dogs (Canis familiaris)’, PLoS ONE, 14(9), p. e0222581. Available at: https://doi.org/10.1371/journal.pone.0222581.
Lerdkrai, C. and Phungphosop, N. (2021) ‘Prevalence of the MDR1 gene mutation in herding dog breeds and Thai Ridgebacks in Thailand’, Veterinary World, 14(11), pp. 3015–3020. Available at: https://doi.org/10.14202/vetworld.2021.3015-3020.
Ley, J. et al. (2007) ‘Assessing fear of novel and startling stimuli in domestic dogs’, Applied Animal Behaviour Science, 104(1), pp. 71–84. Available at: https://doi.org/10.1016/j.applanim.2006.03.021.
Lopez, J.P. et al. (2021) ‘Single-cell molecular profiling of all three components of the HPA axis reveals adrenal ABCB1 as a regulator of stress adaptation’, Science Advances, 7(5), p. eabe4497. Available at: https://doi.org/10.1126/sciadv.abe4497.
Lupien, S.J. et al. (2009) ‘Effects of stress throughout the lifespan on the brain, behaviour and cognition’, Nature Reviews Neuroscience, 10(6), pp. 434–445. Available at: https://doi.org/10.1038/nrn2639.
Lupien, S.J. and McEwen, B.S. (1997) ‘The acute effects of corticosteroids on cognition: integration of animal and human model studies’, Brain Research Reviews, 24(1), pp. 1–27. Available at: https://doi.org/10.1016/S0165-0173(97)00004-0.
Mackin, A.J. et al. (2020) ‘Excessive Cyclosporine-Associated Immunosuppression in a Dog Heterozygous for the MDR1 (ABCB1-1Δ) Mutation’, Journal of the American Animal Hospital Association, 56(3), p. 190. Available at: https://doi.org/10.5326/JAAHA-MS-7004.
Marelli, S.P. et al. (2020) ‘Genotypic and allelic frequencies of MDR1 gene in dogs in Italy’, Veterinary Record Open, 7(1), p. e000375. Available at: https://doi.org/10.1136/vetreco-2019-000375.
Martinez, M. et al. (2008) ‘The pharmacogenomics of P-glycoprotein and its role in veterinary medicine’, Journal of Veterinary Pharmacology and Therapeutics, 31(4), pp. 285–300. Available at: https://doi.org/10.1111/j.1365-2885.2008.00964.x.
Mason, B.L. et al. (2010) ‘Central Nervous System (CNS) Delivery of Glucocorticoids Is Fine-Tuned by Saturable Transporters at the Blood-CNS Barriers and Nonbarrier Regions’, Endocrinology, 151(11), pp. 5294–5305. Available at: https://doi.org/10.1210/en.2010-0554.
Mason, B.L., Pariante, C.M. and Thomas, S.A. (2008) ‘A revised role for P-glycoprotein in the brain distribution of dexamethasone, cortisol, and corticosterone in wild-type and ABCB1A/B-deficient mice’, Endocrinology, 149(10), pp. 5244–5253. Available at: https://doi.org/10.1210/en.2008-0041.
Mason, B.L., Pariante, C.M. and Thomas, S.A. (2012) ‘Changes in the Brain Accumulation of Glucocorticoids in abcb1a-Deficient CF-1 Mice’, Journal of Neuroendocrinology, 24(11), pp. 1440–1446. Available at: https://doi.org/10.1111/j.1365-2826.2012.02353.x.
Maul, S. et al. (2020) ‘Genetics of resilience: Implications from genome-wide association studies and candidate genes of the stress response system in posttraumatic stress disorder and depression’, American Journal of Medical Genetics. Part B, Neuropsychiatric Genetics: The Official Publication of the International Society of Psychiatric Genetics, 183(2), pp. 77–94. Available at: https://doi.org/10.1002/ajmg.b.32763.
McEwen, B.S. (2007) ‘Physiology and Neurobiology of Stress and Adaptation: Central Role of the Brain’, Physiological Reviews, 87(3), pp. 873–904. Available at: https://doi.org/10.1152/physrev.00041.2006.
Mealey, K.L. et al. (2001) ‘Ivermectin sensitivity in collies is associated with a deletion mutation of the mdr1 gene’, Pharmacogenetics, 11(8), pp. 727–733. Available at: https://doi.org/10.1097/00008571-200111000-00012.
Mealey, K.L. et al. (2007) ‘Comparison of the hypothalamic?pituitary?adrenal axis in MDR1-1? and MDR1 wildtype dogs’, Journal of Veterinary Emergency and Critical Care, 17(1), pp. 61–66. Available at: https://doi.org/10.1111/j.1476-4431.2006.00196.x.
Mealey, K. L. et al. (2008) ‘ABCB1-1Delta polymorphism can predict hematologic toxicity in dogs treated with vincristine’, Journal of Veterinary Internal Medicine, 22(4), pp. 996–1000. Available at: https://doi.org/10.1111/j.1939-1676.2008.0122.x.
Mealey, Katrina L. et al. (2008) ‘P-glycoprotein contributes to the blood-brain, but not blood-cerebrospinal fluid, barrier in a spontaneous canine p-glycoprotein knockout model’, Drug Metabolism and Disposition: The Biological Fate of Chemicals, 36(6), pp. 1073–1079. Available at: https://doi.org/10.1124/dmd.107.018978.
Mealey, K.L. et al. (2019) ‘Personalized medicine: going to the dogs?’, Human Genetics, 138(5), pp. 467–481. Available at: https://doi.org/10.1007/s00439-019-02020-w.
Mealey, K.L., Bentjen, S.A. and Waiting, D.K. (2002) ‘Frequency of the mutant MDR1 allele associated with ivermectin sensitivity in a sample population of collies from the northwestern United States’, American Journal of Veterinary Research, 63(4), pp. 479–481. Available at: https://doi.org/10.2460/ajvr.2002.63.479.
Mealey, K.L., Dassanayake, S. and Burke, N.S. (2017) ‘Establishment of a cell line for assessing drugs as canine P-glycoprotein substrates: proof of principle’, Journal of Veterinary Pharmacology and Therapeutics, 40(5), pp. 545–551. Available at: https://doi.org/10.1111/jvp.12390.
Mealey, K.L. and Meurs, K.M. (2008) ‘Breed distribution of the ABCB1-1Δ (multidrug sensitivity) polymorphism among dogs undergoing ABCB1 genotyping’. Available at: https://doi.org/10.2460/javma.233.6.921.
Mealey, K.L., Munyard, K.A. and Bentjen, S.A. (2005) ‘Frequency of the mutant MDR1 allele associated with multidrug sensitivity in a sample of herding breed dogs living in Australia’, Veterinary Parasitology, 131(3–4), pp. 193–196. Available at: https://doi.org/10.1016/j.vetpar.2005.05.004.
Mealey, K.L., Northrup, N.C. and Bentjen, S.A. (2003) ‘Increased toxicity of P-glycoprotein-substrate chemotherapeutic agents in a dog with the MDR1 deletion mutation associated with ivermectin sensitivity’. Available at: https://doi.org/10.2460/javma.2003.223.1453.
Mealey, K.L., Owens, J.G. and Freeman, E. (2023) ‘Canine and feline P‐glycoprotein deficiency: What we know and where we need to go’, Journal of Veterinary Pharmacology and Therapeutics, 46(1), pp. 1–16. Available at: https://doi.org/10.1111/jvp.13102.
Medina-Rodriguez, E.M. and Beurel, E. (2022) ‘Blood brain barrier and inflammation in depression’, Neurobiology of Disease, 175, p. 105926. Available at: https://doi.org/10.1016/j.nbd.2022.105926.
Mills, D., Karagiannis, C. and Zulch, H. (2014) ‘Stress—Its Effects on Health and Behavior’, Veterinary Clinics of North America: Small Animal Practice, 44(3), pp. 525–541. Available at: https://doi.org/10.1016/j.cvsm.2014.01.005.
Müller, M.B. et al. (2003) ‘ABCB1 (MDR1)-Type P-Glycoproteins at the Blood–Brain Barrier Modulate the Activity of the Hypothalamic–Pituitary–Adrenocortical System: Implications for Affective Disorder’, Neuropsychopharmacology, 28(11), pp. 1991–1999. Available at: https://doi.org/10.1038/sj.npp.1300257.
Müller, M.B. et al. (2004) ‘Hypothalamic-Pituitary-Adrenocortical System and Mood Disorders: Highlights from Mutant Mice’, Neuroendocrinology, 79(1), pp. 1–12. Available at: https://doi.org/10.1159/000076041.
Myers, B., McKlveen, J.M. and Herman, J.P. (2014) ‘Glucocorticoid actions on synapses, circuits, and behavior: Implications for the energetics of stress’, Frontiers in Neuroendocrinology, 35(2), pp. 180–196. Available at: https://doi.org/10.1016/j.yfrne.2013.12.003.
Nagasawa, M., Mogi, K. and Kikusui, T. (2012) ‘Continued Distress among Abandoned Dogs in Fukushima’, Scientific Reports, 2(1), p. 724. Available at: https://doi.org/10.1038/srep00724.
Naumovska, Z. et al. (2017) ‘Evaluation of the Role of ABCB1gene Polymorphic Variants on Psychiatric Disorders Predisposition in Macedonian Population’, Prilozi (Makedonska Akademija Na Naukite I Umetnostite. Oddelenie Za Medicinski Nauki), 38(3), pp. 71–88. Available at: https://doi.org/10.2478/prilozi-2018-0008.
Neff, M.W. et al. (2004) ‘Breed distribution and history of canine mdr1-1Δ , a pharmacogenetic mutation that marks the emergence of breeds from the collie lineage’, Proceedings of the National Academy of Sciences, 101(32), pp. 11725–11730. Available at: https://doi.org/10.1073/pnas.0402374101.
de Nooij, L. et al. (2024) ‘Exogenous glucocorticoids to improve extinction learning for post-traumatic stress disorder patients with hypothalamic–pituitary–adrenal-axis dysregulation: a study protocol description’, European Journal of Psychotraumatology, 15(1), p. 2364441. Available at: https://doi.org/10.1080/20008066.2024.2364441.
Notari, L., Burman, O. and Mills, D. (2015) ‘Behavioural changes in dogs treated with corticosteroids’, Physiology & Behavior, 151, pp. 609–616. Available at: https://doi.org/10.1016/j.physbeh.2015.08.041.
Notari, L., Burman, O. and Mills, D.S. (2016) ‘Is there a link between treatments with exogenous corticosteroids and dog behaviour problems?’, Veterinary Record, 179(18), pp. 462–462. Available at: https://doi.org/10.1136/vr.103768.
Notari, L., Kirton, R. and Mills, D.S. (2022) ‘Psycho-Behavioural Changes in Dogs Treated with Corticosteroids: A Clinical Behaviour Perspective’, Animals, 12(5), p. 592. Available at: https://doi.org/10.3390/ani12050592.
Notari, L. and Mills, D. (2011) ‘Possible behavioral effects of exogenous corticosteroids on dog behavior: a preliminary investigation’, Journal of Veterinary Behavior, 6(6), pp. 321–327. Available at: https://doi.org/10.1016/j.jveb.2011.02.004.
Nürnberger, D. et al. (2021) ‘First Sequencing of Caprine Mdr1 (Abcb1) mRNA Due to Suspected Neurological Adverse Drug Reaction in a Thuringian Goat Following Extra-Label Use of Doramectin’, Frontiers in Veterinary Science, 8. Available at: https://doi.org/10.3389/fvets.2021.682393.
Orzechowski, K.L. et al. (2012) ‘Neurotoxic effects of ivermectin administration in genetically engineered mice with targeted insertion of the mutated canine ABCB1 gene’. Available at: https://doi.org/10.2460/ajvr.73.9.1477.
Pan, X. et al. (2018) ‘Salivary cortisol in post-traumatic stress disorder: a systematic review and meta-analysis’, BMC Psychiatry, 18(1), p. 324. Available at: https://doi.org/10.1186/s12888-018-1910-9.
Papilloud, A. et al. (2019) ‘Peripubertal stress-induced heightened aggression: modulation of the glucocorticoid receptor in the central amygdala and normalization by mifepristone treatment.’, Neuropsychopharmacology, 44(4), pp. 674–682. Available at: https://doi.org/10.1038/s41386-018-0110-0.
Pekcec, A. et al. (2011) ‘Age-dependent decline of blood–brain barrier P-glycoprotein expression in the canine brain’, Neurobiology of Aging, 32(8), pp. 1477–1485. Available at: https://doi.org/10.1016/j.neurobiolaging.2009.08.014.
Petryszyn, P.W. and Wiela-Hojeńska, A. (2018) ‘The importance of the polymorphisms of the ABCB1 gene in disease susceptibility, behavior and response to treatment in inflammatory bowel disease: A literature review’, Advances in Clinical and Experimental Medicine: Official Organ Wroclaw Medical University, 27(10), pp. 1459–1463. Available at: https://doi.org/10.17219/acem/92936.
Pigott, A. (2016) ‘Pain management’, in R. Kirby and A. Linklater (eds) Monitoring and Intervention for the Critically Ill Small Animal. 1st edn. Wiley, pp. 333–348. Available at: https://doi.org/10.1002/9781118923870.ch19.
Qing Yang et al. (2015) ‘Contribution of MDR1 gene polymorphisms on IBD predisposition and response to glucocorticoids in IBD in a Chinese population.’, Journal of Digestive Diseases, 16(1), pp. 22–30. Available at: https://doi.org/10.1111/1751-2980.12205.
Radosta, L. (2024) ‘Behavior Changes Associated with Metabolic Disease of Dogs and Cats’, Veterinary Clinics: Small Animal Practice, 54(1), pp. 17–28. Available at: https://doi.org/10.1016/j.cvsm.2023.08.004.
Raison, C.L. and Miller, A.H. (2003) ‘When Not Enough Is Too Much: The Role of Insufficient Glucocorticoid Signaling in the Pathophysiology of Stress-Related Disorders’, American Journal of Psychiatry, 160(9), pp. 1554–1565. Available at: https://doi.org/10.1176/appi.ajp.160.9.1554.
Reul, J.M.H.M. et al. (1990) ‘Binding Characteristics of Mineralocorticoid and Glucocorticoid Receptors in Dog Brain and Pituitary’, Endocrinology, 127(2), pp. 907–915. Available at: https://doi.org/10.1210/endo-127-2-907.
Richter-Levin, G., Stork, O. and Schmidt, M.V. (2019) ‘Animal models of PTSD: a challenge to be met’, Molecular Psychiatry, 24(8), pp. 1135–1156. Available at: https://doi.org/10.1038/s41380-018-0272-5.
Rosado, B. et al. (2010) ‘The role of the hypothalamic-pituitary-adrenal axis in canine aggression towards humans’, Journal of Veterinary Behavior, 5(1), pp. 29–30. Available at: https://doi.org/10.1016/j.jveb.2009.10.030.
Roulet, A. et al. (2003) ‘MDR1-deficient genotype in Collie dogs hypersensitive to the P-glycoprotein substrate ivermectin’, European Journal of Pharmacology, 460(2–3), pp. 85–91. Available at: https://doi.org/10.1016/s0014-2999(02)02955-2.
Salden, S. et al. (2023) ‘Disaster response and its aftermath: A systematic review of the impact of disaster deployment on working dogs’, Applied Animal Behaviour Science, 265, p. 105987. Available at: https://doi.org/10.1016/j.applanim.2023.105987.
Salonen, M. et al. (2020) ‘Prevalence, comorbidity, and breed differences in canine anxiety in 13,700 Finnish pet dogs’, Scientific Reports, 10(1), p. 2962. Available at: https://doi.org/10.1038/s41598-020-59837-z.
Salvatore, J.E. et al. (2015) ‘Genome-wide association data suggest ABCB1 and immune-related gene sets may be involved in adult antisocial behavior’, Translational Psychiatry, 5(4), pp. e558–e558. Available at: https://doi.org/10.1038/tp.2015.36.
Santos, M. et al. (2014) ‘Common Genetic Polymorphisms in the ABCB1 Gene Are Associated with Risk of Major Depressive Disorder in Male Portuguese Individuals’, Genetic Testing and Molecular Biomarkers, 18(1), pp. 12–19. Available at: https://doi.org/10.1089/gtmb.2013.0197.
Savas, M. et al. (2020) ‘Systemic and Local Corticosteroid Use Is Associated with Reduced Executive Cognition, and Mood and Anxiety Disorders’, Neuroendocrinology, 110(3–4), pp. 282–291. Available at: https://doi.org/10.1159/000501617.
Schinkel, A.H. et al. (1994) ‘Disruption of the mouse mdr1a P-glycoprotein gene leads to a deficiency in the blood-brain barrier and to increased sensitivity to drugs’, Cell, 77(4), pp. 491–502. Available at: https://doi.org/10.1016/0092-8674(94)90212-7.
Schinkel, A.H. et al. (1997) ‘Normal viability and altered pharmacokinetics in mice lacking mdr1-type (drug-transporting) P-glycoproteins’, Proceedings of the National Academy of Sciences of the United States of America, 94(8), pp. 4028–4033. Available at: https://doi.org/10.1073/pnas.94.8.4028.
Schmidt, M.V. and Müller, M.B. (2006) ‘Animal models of anxiety’, Drug Discovery Today: Disease Models, 3(4), pp. 369–374. Available at: https://doi.org/10.1016/j.ddmod.2006.11.002.
Schoenfelder, Y., Hiemke, C. and Schmitt, U. (2012) ‘Behavioural Consequences of P-Glycoprotein Deficiency in Mice, with Special Focus on Stress-Related Mechanisms’, Journal of Neuroendocrinology, 24(5), pp. 809–817. Available at: https://doi.org/10.1111/j.1365-2826.2012.02278.x.
Seah, C. et al. (2022) ‘Modeling gene × environment interactions in PTSD using human neurons reveals diagnosis-specific glucocorticoid-induced gene expression’, Nature Neuroscience, 25(11), pp. 1434–1445. Available at: https://doi.org/10.1038/s41593-022-01161-y.
Segurson, S. and Smith-Blackmore, M. (2024) ‘Preparing veterinarians to respond to animal victims: understanding and treating the behavioral consequences of maltreatment’, in L. Levitt, D.B. Rosengard, and J. Rubin (eds) Animals as Crime Victims. Edward Elgar Publishing, pp. 254–267. Available at: https://doi.org/10.4337/9781802209884.00025.
Sep, M.S.C., Geuze, E. and Joëls, M. (2023) ‘Impaired learning, memory, and extinction in posttraumatic stress disorder: translational meta-analysis of clinical and preclinical studies’, Translational Psychiatry, 13(1), pp. 1–9. Available at: https://doi.org/10.1038/s41398-023-02660-7.
Sherman, B.L. et al. (2015) ‘A test for the evaluation of emotional reactivity in Labrador retrievers used for explosives detection’, Journal of Veterinary Behavior, 10(2), pp. 94–102. Available at: https://doi.org/10.1016/j.jveb.2014.12.007.
Spaulding, K. (2022) The stress factor in dogs: unlocking resiliency and enhancing well-being. Wenatchee, Washington: Dogwise Publishing.
Stafford, E.G. et al. (2021) ‘PREVALENCE OF THE ABCB1-1Δ GENE IN NONDOMESTIC SPECIES OF THE CANIDAE FAMILY’, Journal of Zoo and Wildlife Medicine, 51(4), pp. 1007–1011. Available at: https://doi.org/10.1638/2020-0057.
Stampfl, B. (2012) ‘Theorizing Canine PTSD’:, in Semiotics. Semiotic Society of America, pp. 159–168. Available at: https://doi.org/10.5840/cpsem201216.
Stuart, F.A., Segal, T.Y. and Keady, S. (2005) ‘Adverse psychological effects of corticosteroids in children and adolescents’, Archives of Disease in Childhood, 90(5), pp. 500–506. Available at: https://doi.org/10.1136/adc.2003.041541.
Swain, M.D. et al. (2013) ‘P-gp substrate-induced neurotoxicity in an Abcb1a knock-in/Abcb1b knock-out mouse model with a mutated canine ABCB1 targeted insertion’, Research in Veterinary Science, 94(3), pp. 656–661. Available at: https://doi.org/10.1016/j.rvsc.2012.10.025.
Szeszko, P.R., Lehrner, A. and Yehuda, R. (2018) ‘Glucocorticoids and Hippocampal Structure and Function in PTSD’, Harvard Review of Psychiatry, 26(3), p. 142. Available at: https://doi.org/10.1097/HRP.0000000000000188.
Tappin, S.W. et al. (2012) ‘Frequency of the mutant MDR1 allele in dogs in the UK’, Veterinary Record, 171(3), pp. 72–72. Available at: https://doi.org/10.1136/vr.100633.
Thoeringer, C.K. et al. (2007) ‘Multidrug-resistance gene 1-type p-glycoprotein (MDR1 p-gp) inhibition by tariquidar impacts on neuroendocrine and behavioral processing of stress’, Psychoneuroendocrinology, 32(8–10), pp. 1028–1040. Available at: https://doi.org/10.1016/j.psyneuen.2007.08.001.
Tseilikman, V. et al. (2021) ‘A Rat Model of Post-Traumatic Stress Syndrome Causes Phenotype-Associated Morphological Changes and Hypofunction of the Adrenal Gland’, International Journal of Molecular Sciences, 22(24), p. 13235. Available at: https://doi.org/10.3390/ijms222413235.
Uhr, M., Holsboer, F. and Müller, M.B. (2002) ‘Penetration of endogenous steroid hormones corticosterone, cortisol, aldosterone and progesterone into the brain is enhanced in mice deficient for both mdr1a and mdr1b P-glycoproteins’, Journal of Neuroendocrinology, 14(9), pp. 753–759. Available at: https://doi.org/10.1046/j.1365-2826.2002.00836.x.
Ullmann, E. et al. (2020) ‘Offensive Behavior, Striatal Glutamate Metabolites, and Limbic–Hypothalamic–Pituitary–Adrenal Responses to Stress in Chronic Anxiety’, International Journal of Molecular Sciences, 21(20), p. 7440. Available at: https://doi.org/10.3390/ijms21207440.
Ulrich-Lai, Y.M., James P. Herman and Herman, J.P. (2009) ‘Neural regulation of endocrine and autonomic stress responses.’, Nature Reviews Neuroscience, 10(6), pp. 397–409. Available at: https://doi.org/10.1038/nrn2647.
Varghese, S.M. et al. (2024) ‘Unraveling the Role of the Blood-Brain Barrier in the Pathophysiology of Depression: Recent Advances and Future Perspectives’, Molecular Neurobiology [Preprint]. Available at: https://doi.org/10.1007/s12035-024-04205-5.
Verbitsky, A. et al. (2020) ‘Rodent models of post-traumatic stress disorder: behavioral assessment.’, Translational Psychiatry, 10(1), pp. 132–132. Available at: https://doi.org/10.1038/s41398-020-0806-x.
Wormald, D. et al. (2016) ‘Physiological stress coping and anxiety in greyhounds displaying inter-dog aggression’, Applied Animal Behaviour Science, 180, pp. 93–99. Available at: https://doi.org/10.1016/j.applanim.2016.04.007.
Wright, H.F., Mills, D.S. and Pollux, P.M.J. (2011) ‘Development and Validation of a Psychometric Tool forAssessing Impulsivity in the Domestic Dog ( Canis familiaris)’, International Journal of Comparative Psychology, 24(2). Available at: https://doi.org/10.46867/IJCP.2011.24.02.03.
Yau, J.L.W. et al. (2007) ‘The Antidepressant Desipramine Requires the ABCB1 (Mdr1)-Type p-Glycoprotein to Upregulate the Glucocorticoid Receptor in Mice’, Neuropsychopharmacology, 32(12), pp. 2520–2529. Available at: https://doi.org/10.1038/sj.npp.1301389.
Yehuda, R. (2002) ‘Post-Traumatic Stress Disorder’, New England Journal of Medicine, 346(2), pp. 108–114. Available at: https://doi.org/10.1056/NEJMra012941.
Yehuda, R. (2009) ‘Status of glucocorticoid alterations in post-traumatic stress disorder.’, Annals of the New York Academy of Sciences, 1179(1), pp. 56–69. Available at: https://doi.org/10.1111/j.1749-6632.2009.04979.x.
Yehuda, R. et al. (2015) ‘Post-traumatic stress disorder’, Nature Reviews Disease Primers, 1(1), p. 15057. Available at: https://doi.org/10.1038/nrdp.2015.57.
Yehuda, R. and Seckl, J. (2011) ‘Minireview: Stress-Related Psychiatric Disorders with Low Cortisol Levels: A Metabolic Hypothesis’, Endocrinology, 152(12), pp. 4496–4503. Available at: https://doi.org/10.1210/en.2011-1218.
Zanette, L.Y. et al. (2019) ‘Predator-induced fear causes PTSD-like changes in the brains and behaviour of wild animals’, Scientific Reports, 9(1), p. 11474. Available at: https://doi.org/10.1038/s41598-019-47684-6.